Endoplazmik retikulum (kelimenin tam anlamıyla ‘ plazma ağının içinde’ ), hücredeki ana sentez bölgesidir.
Dış nükleer zarf ile uzayan (ve sürekli olan) bir düzleştirilmiş keseler sistemi (sarnıç) sistemidir – hücrenin önemli bir hacmini alır (hücredeki membranöz bileşenlerin yarısından fazlası). Fizyolojik işlevi Golgi cihazınınki ile çok yakın bir ilişkiye sahiptir ve birlikte hücrenin salgı yolunu oluştururlar.
‘Endoplazmik retikulum’, özel dokuda boyut (örneğin GI kanalındaki salgı hücreleri) ve fonksiyonu (örneğin kasta sarkoplazmik retikulum) gibi küçük değişikliklerle granüllü veya granülsüz olarak sınıflandırılır. Sonuçta, bunlar ultra yapısal olarak farklılık gösterir ve bu, sınıflandırmalarının temelidir.
Yazı Başlıkları
Granüllü Endoplazmik Retikulum (RER)
Granüllü (rough) endoplazmik retikulum, sitoplazmik yüzeye bağlı birçok ribozom içerir ve bunlar, hedeflenen proteinlerin sentezinde önemli bir rol oynar:
- Hücre dışı matrikse salınım (ECM) (örn. Mukus ve enzimler)
- Plasmalemma ile birleşme (örn. Reseptörler ve kanallar)
- Membran bağlı veziküller (örneğin lizozom enzimleri)
Bunun gerçekleşmesi için gereken tüm mekanizmalar, Golgi cihazı tarafından tamamlanan protein hedeflemesinin son aşamaları ile birlikte, protein hedeflemesinin ilk aşamaları da dahil olmak üzere, RER’de gerçekleşir. Sitosol için hedeflenen proteinler, başlangıçta, sitoplazmada askıya alınmış serbest ribozomlar tarafından kopyalanır.
RER’in bu kadar tanımlayıcı bir özelliği olan ribozomların varlığına rağmen, RER-ribozomal etkileşimleri kalıcı değildir ve bu nedenle protein salgılanması talebine bağlı olarak bağlanma ve ayrılma sürelerinden geçer. Yüzeye yapışma enerjiye bağlıdır ve bu yüzden ATP / GTP sentezi azaldığında, hipoksik hücre hasarında ribozomların koptuğunu görüyoruz.
RER lümenine çevrilen proteinler daha sonra bir dizi post-translasyonel modifikasyona tabi tutulur. Pek çok salgılayıcı protein üreten hücreler, RER ve mitokondriyi genişletecektir.
Çalışmadan OpenStax [CC BY 4.0 (https://creativecommons.org/licenses/by/4.0)] tarafından Wikimedia Commons aracılığıyla uyarlanmıştır.
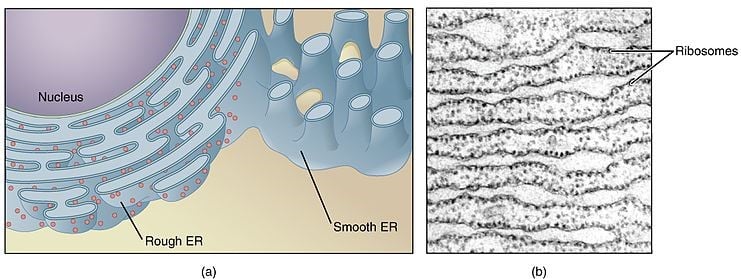
Şekil 1 – Şema gösterimi a) Granüllü endoplazmik retikulum ile düz endoplazmik retikulum arasındaki yapıdaki fark ve b) Granüllü endoplazmik retikulumun elektron mikroskobu
Salgı Yolunun Endoplazmik Kısmı
Salgı yolunun üç ana organelleri vardır: çekirdek, ER ve golgi aparatı. Yolun ER kısmına yoğunlaşarak, konuma göre iki aşamaya ayrılabilir:
Sitoplazmada
- mRNA kopyalanır ve çekirdekten nükleer gözenekler aracılığıyla çıkar. Sitoplazmada serbest ribozomlarla birleşirler.
- Üretilen tüm proteinler başlangıçta, sitozolde serbest şekilde askıya alınan ribozomlar yoluyla translasyona başlar. Bu proteinleri RER’e hedefleyen (ve böylece salgılama), sinyal dizisini kodlayan mRNA’nın iç dizisidir.
- Yeni (yeni oluşmuş) proteinin sinyal sekansı, serbest ribozom (peptid-ribozom kompleksi) tarafından çevrildikten sonra, başka bir önemli protein, bunu tanır ve kompleksi RER’e yönlendirmekle sorumludur, sinyal peptit (SRP) . SRP ayrıca sitoplazmada serbesttir.
- SRP’nin bağlanması çeviriyi durdurur. Bu, düzenleyici bir işleve hizmet ettiği için önemlidir. RER’de sadece SRP için ayrık sayıda bağlanma bölgesi bulunduğundan, hedef konumlanıncaya kadar sentezi önlemek için sentezin kontrol edilmesi gerekir. Bu, tüm bağlanma bölgeleri işgal edildiğinde aşırı proteinlerin tamamen sitoplazmaya sentezlenmesini önler.
- Peptit-ribozom bağlı SRP de GTP molekülüne bağlanır. Bu, RER ile ribozomal ilişkinin enerjiye bağımlı bileşenidir. GTP, sentezin yeniden aktifleştirilmesi için önemlidir.
Yukarıda tartışıldığı gibi, GTP tedarikinin azaltılması, ribozomların bağlanmasını önler ve salgılayıcı protein sentezinin yukarıda tartışıldığı gibi gerçekleşmesini sağlar. GTP, Süksinil-CoA’dan TCA döngüsü yoluyla sentezlenir.
GTP-SRP’ye bağlı peptid-ribozom kompleksi, daha sonra, SRP reseptörünün (aka kenetlenme proteini) bulunduğu granüllü endoplazmik membrana doğru lokalize edilir. SRP reseptörü, yeni ortaya çıkan proteinin RER’e girmesini sağlayan bir kanala bitişiktir. Bu, translocon. SRP’nin SRP reseptörü ile bağlanması GTP hidrolizine yol açar.
Translocon açılır ve SRP ayrıştırılır. Ortaya çıkan proteinin sentezi bu translokona ve dolayısıyla RER’ye devam edebilir. Salgılayıcı bir protein ise, proteinin RER içine hareketi tamamlanır. Transmembran proteinlerinde, translasyonları durdurulur ve lateral bir geçiş mekanizması, translokondan proteini zarın içine bırakır.
Sinyal sekansı zaten çevrilmiş olduğundan, önce translokon üzerinden RER’ye girer.
Bir enzim, sinyal peptidazı , sinyal dizisini ayırır ve böylece proteini salgılama yoluna iletir.
RER’de Çeviri Sonrası Değişiklik
Tüm protein RER’e girdiğinde, uygun işleyişi ve taşınması için modifikasyonlar yapılır.
Değişiklikler şunları içerir:
- Proteolitik bölünme (inhibitör alanların çıkarılmasıyla translasyon sonrası modifikasyon).
- Üçüncü katlamanın sağlanması için disülfür bağ oluşumu.
- Amino asidin hidroksilasyonu kalır (glikozilasyona yardımcı olur).
- N bağlantılı glikosilasyon – azot artıkları üzerine karbonhidrat ilavesi. Diğer moleküllerle etkileşimi kolaylaştırmak önemlidir (örneğin hücresel adezyon).
Protein modifiye edildikten sonra, bir vezikül içine sıkışır ve Golgi aparatına doğru hareket eder, burada vücudun etrafını hedef almak için daha fazla modifiye edilir ve paketlenir.
Serbest ribozomlar tarafından üretilen proteinler nadiren glikosile edilir.
Granülsüz Endoplazmik Retikulum (SER)
Granülsüz (Smooth) endoplazmik retikulum, lipitler, fosfolipitler ve steroidlerin sentezinde önemlidir. Genellikle daha az yaygındır ve bazı özel dokularda (örn. Steroidojenik hücreler ve kaslar) sıklıkla SER genişlemesine sahip olmasına rağmen ribozomlar bununla ilişkilendirilmez.
Aynı zamanda alkol ve barbitüratlar gibi bazı ilaçların ve toksinlerin (ksenobiyotikler) detoksikasyonunda önemli olan sitokrom P450 enzimlerini de içerirler. Hepatositler, SER bakımından zengin bölgelerde glikojeni depolar.
Kas içerisinde bulunan granülsüz endoplazmik retikulum sarkoplazmik retikulum olarak bilinir ve özel bir fonksiyona hizmet eder.
Çalışmadan OpenStax [CC BY 4.0 (https://creativecommons.org/licenses/by/4.0)] tarafından Wikimedia Commons aracılığıyla uyarlanmıştır.
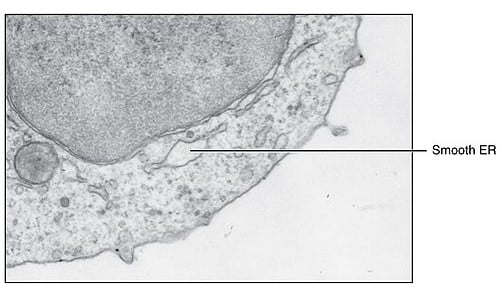
Şekil 2 – Granülsüz Endoplazmik Retikulumun Elektron Mikroskobu
Sarkoplazmik retikulum (SR)
Kastaki sarkoplazmik retikulum geniştir, çünkü kasılma (ve gevşeme) tonik kuvvetini modüle eden kalsiyum sekestrasyonunda önemli bir rol oynar.
Sarkoplazmik retikulumun üst yapısı kas tipleri arasında farklılık gösterir. Çizgili kasta, dik T tübülünün etrafına yerleştirilmişlerdir, burada eylem potansiyeli / yüzey Ca2 + girişi kalsiyum yükselişini başlatabilir. Bu, diadlere (kalp kası) ve triadlara (iskelet kası) yol açar. Farklı kas tipleri, farklı SR ekstansiyonlarına sahiptir.
Klinik: Malign Hipertermi
Malign hipertermi (MH), kastaki sarkoplazmik retikulumun ryanodin reseptörünün mutasyonuyla sonuçlanan akut bir durumdur. Mutasyon, alıcının herhangi bir potansiyel agoniste duyarlılığının artmasına neden olur ve abartılı bir kalsiyum salınımına yol açar, bunun sonucunda hipermetabolik kriz olarak bilinir – yüksek miktarda oksijen tüketimi ile kas metabolizmasının hızlı bir şekilde hızlandırılması.
Ölüme neden olan faktör, SERCA tarafından Ca2 + ‘ nın büzülmesi ve tutulması için hızlandırılmış oksidatif fosforilasyona eşlik eden vücut sıcaklığının hızlı yükselmesidir, böylece vücudun telafi etme kabiliyetini aşar. Tükenmiş ATP, fosfat, kreatin kinaz, miyoglobin ve potasyum, görülen krampa ve sonrasında kas hasarına katkıda bulunur. Asidozun bir sonucu olarak sistemik etkiler organ fonksiyon bozukluğuna neden olabilir
MH için en yaygın çöktürme faktörü, süksinilkolin gibi uçucu anestezik ajanların kullanılmasıdır . Kas gevşeticiler – dantrolen ve tetikleyicinin kesilmesi tercih edilen tedavilerdir. Dantrolen, SR’den Ca2 + salınımını inhibe ederek çalışır.